023 — PING fundamentals
Abstract
Strips PING down to its biophysical fundamentals and characterises it in isolation from any task — no training, no readout, no loss, just a free-running network driven by Poisson input. The E → I → E loop produces gamma at ≈30 Hz with the standard cycle period, and the f-I curves show the loop is dynamic-range compression: PING holds E an order of magnitude below COBA across two orders of magnitude of drive, while COBA saturates near its refractory ceiling.
Methods
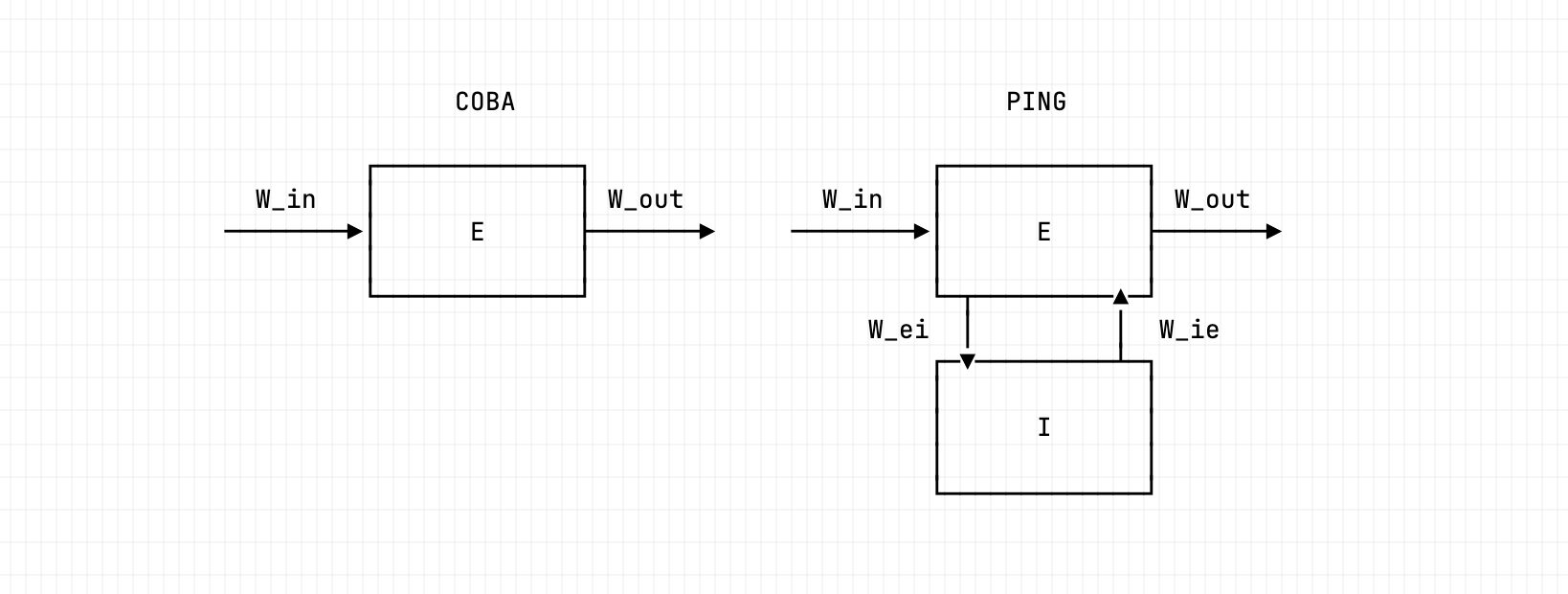
The PING architecture characterised in this notebook. An input layer drives the excitatory population through ; the E→I→E loop is formed by (E to I) and (I back to E), with no I→I synapse. Disabling the I→E loop (ei-strength 0) recovers the COBA reference; engaging it (ei-strength 1.5) produces the gamma rhythm.
The full conductance-based model that produces PING is documented in COBANet; this notebook is the empirical companion piece. Two conditions are run on the same network: ei-strength 0 (loop disabled, the COBA reference) and ei-strength 1.5 (loop engaged, the PING regime). MNIST digit 0 drives the input layer through trained . The rate sweep at the end replays the trained PING baseline from nb025 (seed 42, off) over a sweep of input rates, with both MNIST and spatially uniform Poisson drive.
Results
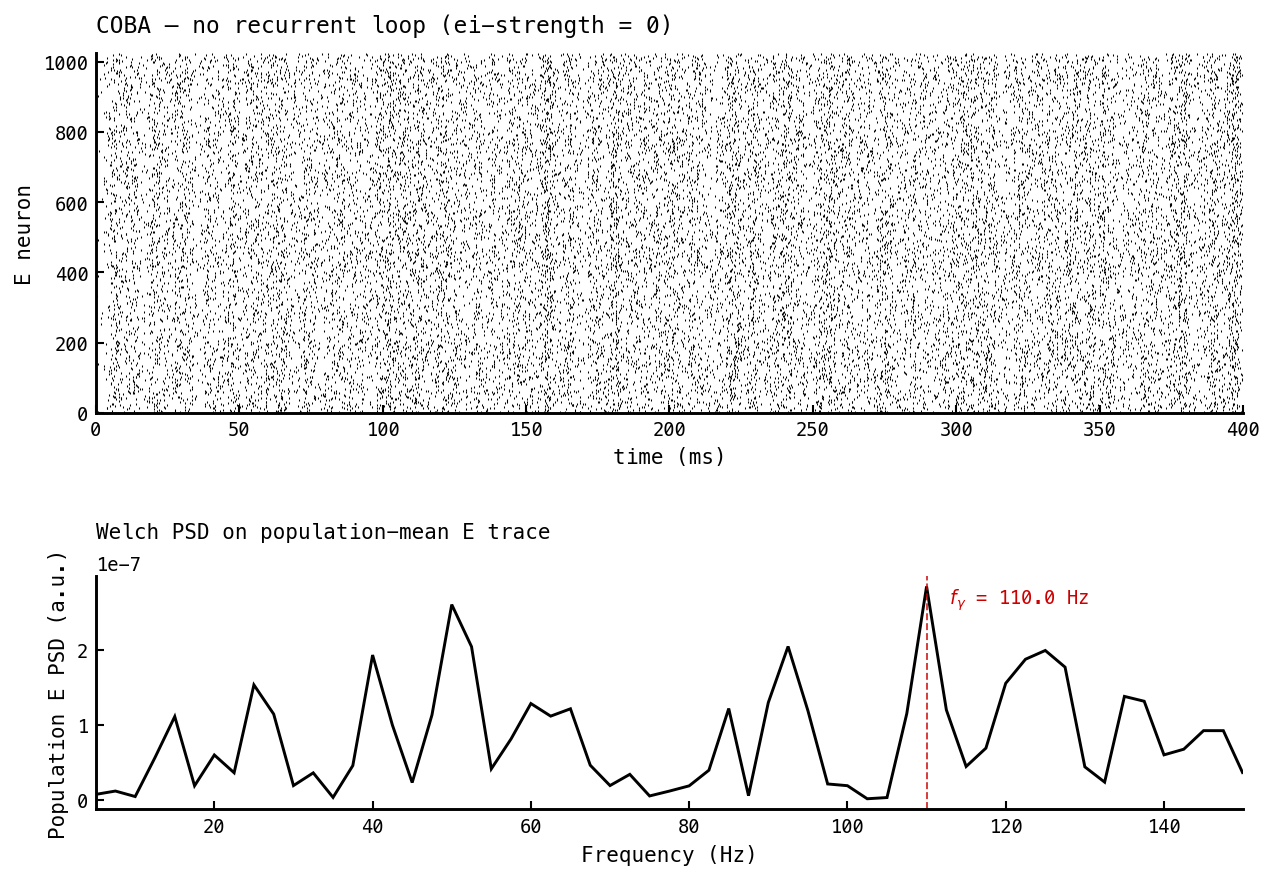
Baseline: recurrent inhibitory loop silent (—ei-strength 0). The E raster fires asynchronously across the full 400 ms; the I population is silent. The Welch PSD on the population-mean E trace (computed exactly like nb041 and nb049: one window per trial, nperseg = T, parabolic-interpolated peak) shows no isolated gamma peak — only the broadband structure of input-driven asynchronous firing. This is the “PING off” reference for the next figure.
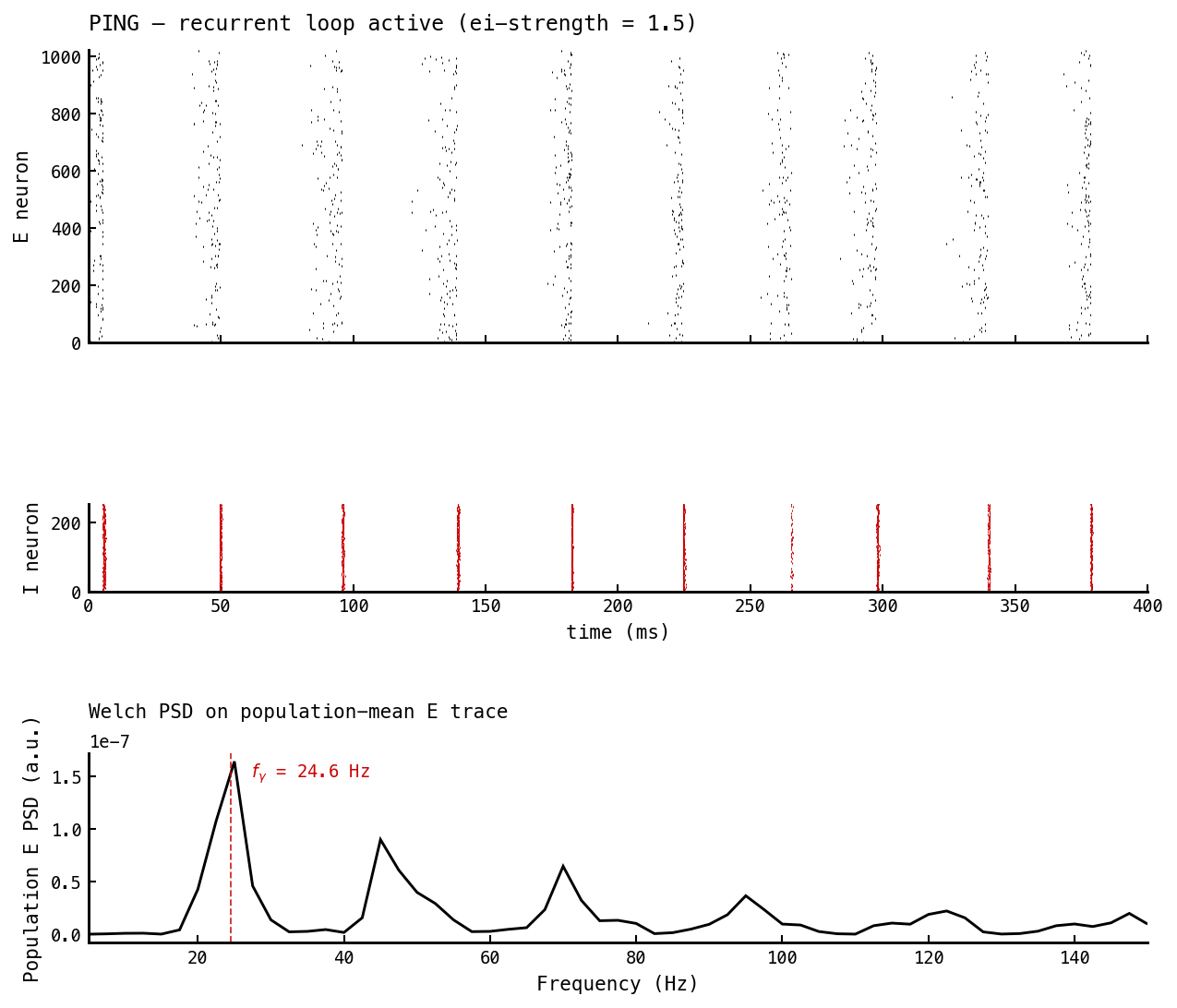
Same network with the I→E loop turned on (—ei-strength 1.5). Same MNIST input, same integration window. The E raster now shows vertical bands and the I population fires synchronous bursts trailing each E-burst by the AMPA delay. The Welch PSD has a clean gamma peak at Hz, with the periodic burst structure visible as harmonics at the higher integer multiples — the spectral signature of the recurrent E↔I loop that the COBA panel lacked.
COBA single-neuron traces. Three panels for the most active E neuron — voltage, conductances, signed currents — with no I-loop activity to show.
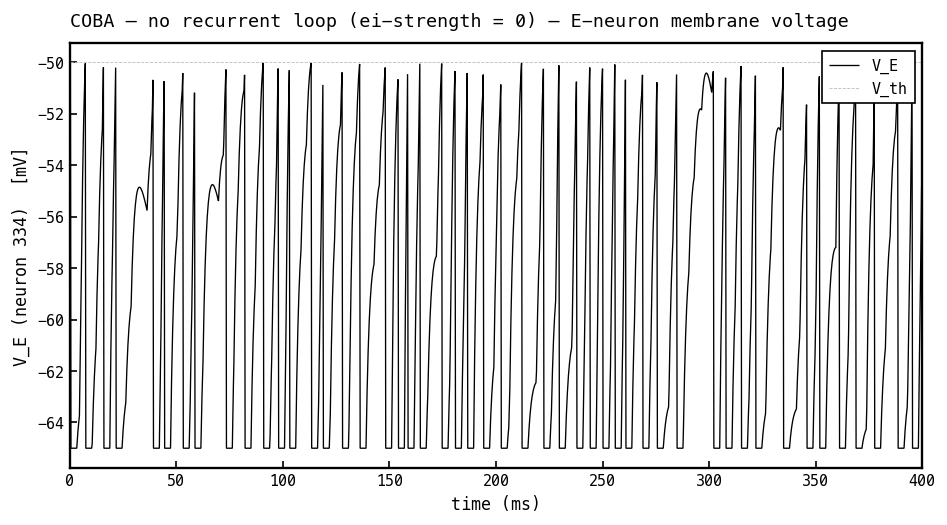
Membrane voltage (black) of the most active E cell over 400 ms with the recurrent loop off (COBA mode, ei-strength 0). Driven only by feedforward input, the cell just charges toward threshold ( mV, faint dashed) and hard-resets to mV each time it fires — irregular, input-paced spiking with no rhythmic structure. This is the rhythm-free control for the PING version in Figure 4a, where inhibition carves a gamma envelope into the same trace.
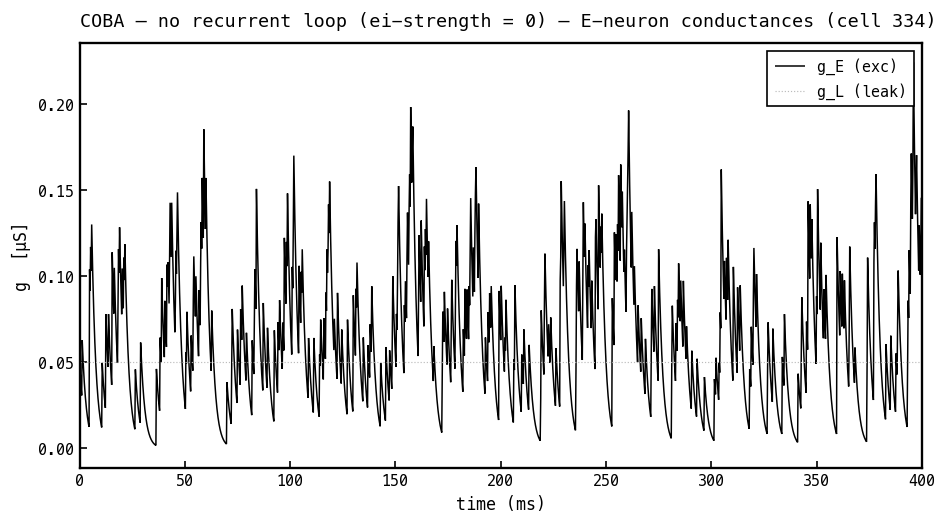
Synaptic and leak conductances on the same E cell, COBA mode. Only the excitatory conductance (black) is active — it steps up with each arriving input spike and decays with — while the leak (faint dotted) is a fixed constant. There is no inhibitory because the I→E loop is off. All conductances are non-negative: they count open channels, not current direction or sign.
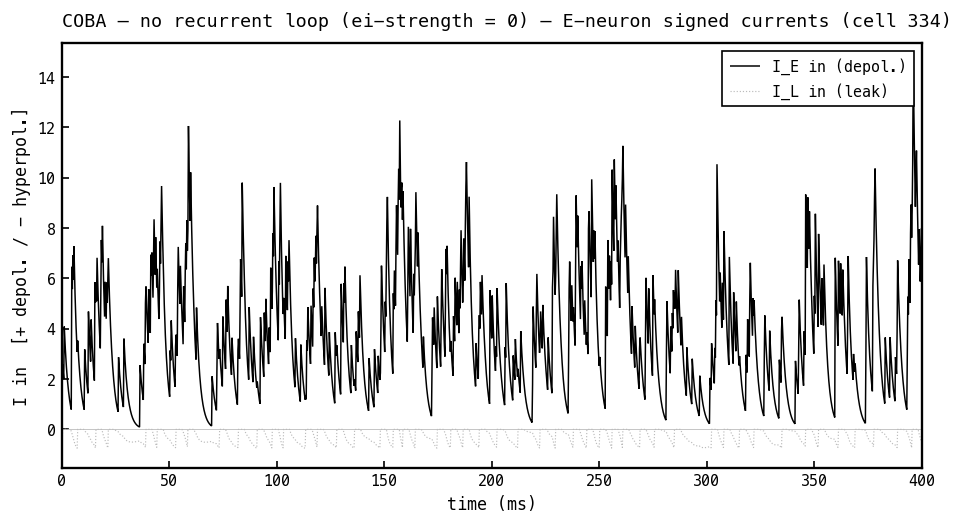
Signed synaptic currents into the E cell, COBA mode, with (positive = depolarising, i.e. pushes up). With no inhibition, the only synaptic current is the depolarising excitatory current (black); the leak current (faint) hovers near zero. Compare Figure 4c, where adding the loop introduces an inhibitory current of the opposite sign.
PING single-neuron traces. Six panels: three for the most active E neuron, three for the most active I neuron. The E panels are where the “sign lives in the driving force” picture becomes literal.
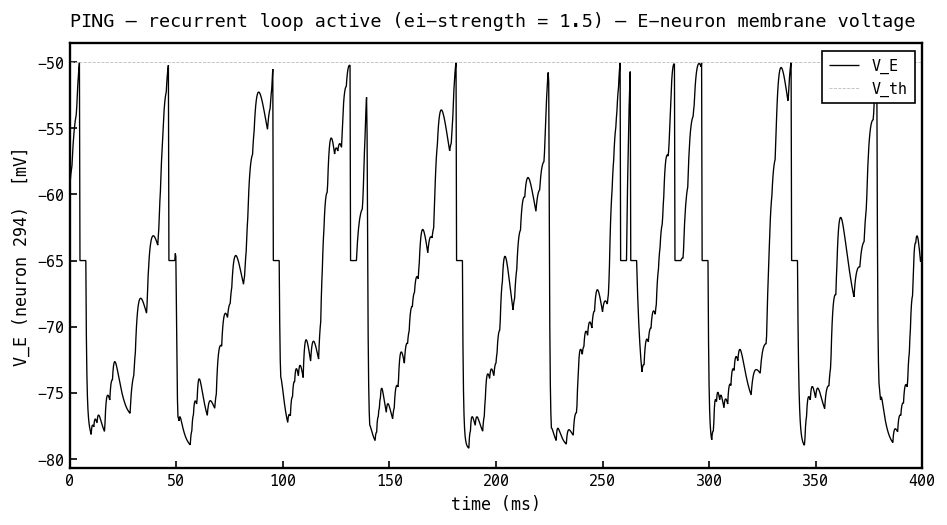
Membrane voltage (black) of the most active E cell with the recurrent loop on (PING mode, ei-strength 1.5), same input as the COBA panel 3a. Now the rhythmic inhibitory bursts hold the cell below threshold most of the time: each I-burst drags down toward mV, and the membrane recovers — and can fire — only as the inhibition decays between bursts. The gamma rhythm is visible directly in the voltage; the loop has turned the irregular firing of 3a into cycle-gated firing.
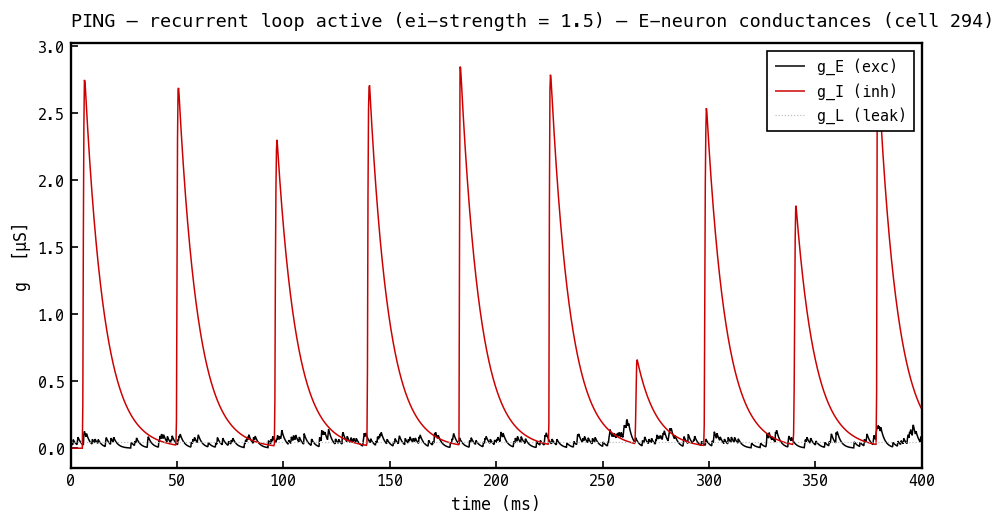
Conductances on the same E cell, PING mode. The inhibitory conductance (red) now dominates — it spikes once per gamma cycle as the synchronous I-burst arrives through , then decays with — while the excitatory (black) carries the smaller feedforward input and (faint) is fixed. All three traces are non-negative: is large and positive, and that is what shunts the cell — it carries no sign of its own.
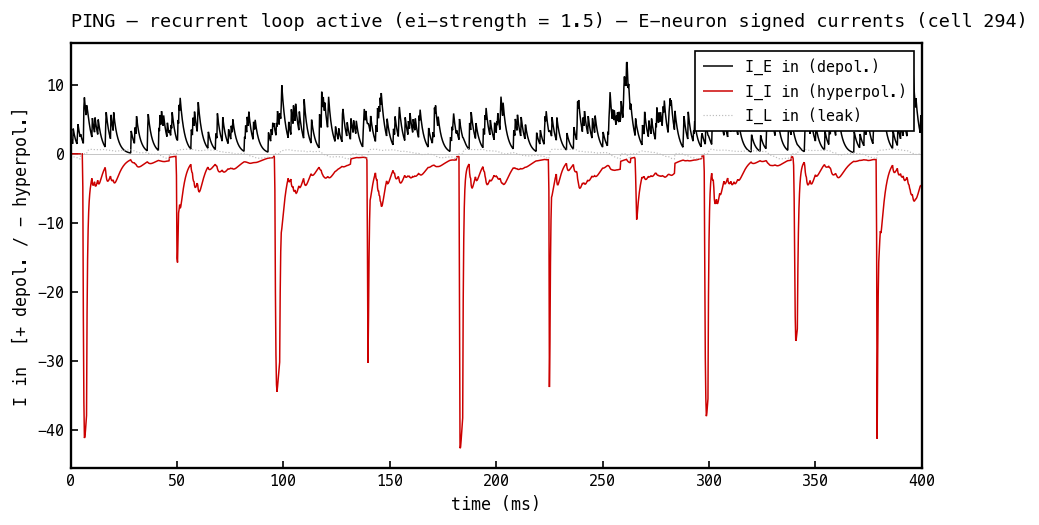
The load-bearing panel. Signed currents into the E cell, PING mode. The excitatory current (black) is positive (depolarising). The inhibitory current (red) is negative (hyperpolarising) even though is positive (Figure 4b) — the minus sign comes entirely from the driving force whenever sits above mV. This is the COBA principle made literal: the sign lives in the driving force, never in the conductance. The sharp negative pulses are the per-cycle inhibitory kicks that gate the rhythm.
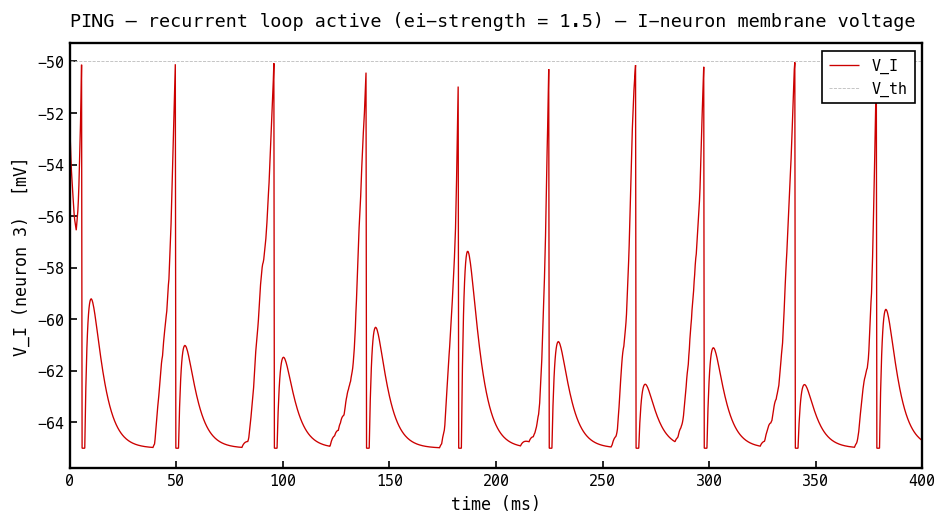
Membrane voltage (red) of the most active I cell, PING mode. The I cell integrates the excitatory volley from the E population and fires once per gamma cycle, so tracks the E-burst envelope — ramping up as E cells fire, crossing threshold ( mV, faint dashed), resetting, and waiting for the next volley. This single I-spike per cycle is what delivers the inhibitory burst seen on the E cell in 4b–4c.
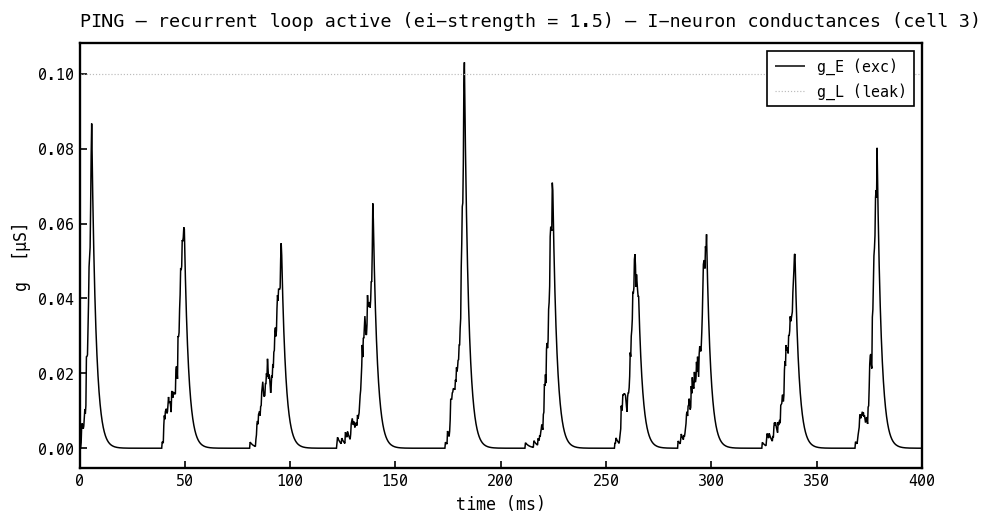
Conductances on the I cell, PING mode. The I cell receives only excitation: (black) is the arriving E-spike envelope (delivered via ) and (faint) is the fixed leak. There is no inhibitory conductance — the architecture has no I→I synapse — which is exactly why the I population can synchronise into the sharp once-per-cycle bursts that clock the loop.
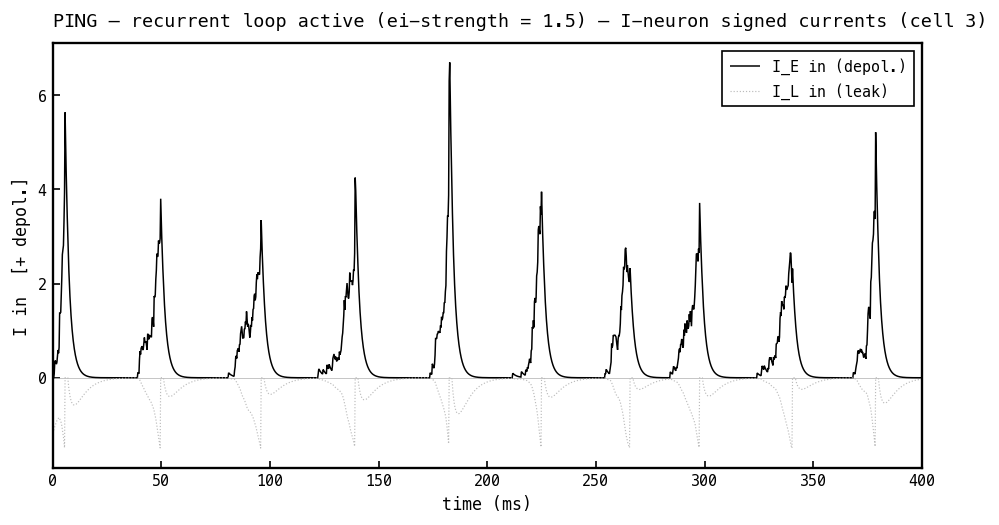
Signed currents into the I cell, PING mode. With no inhibitory input, only the depolarising excitatory current (black) and a small leak current contribute — the I cell is driven purely up to threshold. Read 4a–4f together and the loop is in cross-section: E spikes ramp on I (4e–4f) → I fires once per cycle (4d) → that spike delivers the inhibitory burst on E (4b) → its negative current shuts E down (4c, 4a) → decays → E refires. That delayed E→I→E loop is PING.
Input-rate sweep
The rasters and traces above are at a single working point. Sweeping the Poisson input rate across the trained PING baseline (nb025, seed 42, off) shows how the network responds to drive: silent below a recruitment threshold, gamma-locked above it, with cycle period invariant to further drive once the loop engages.
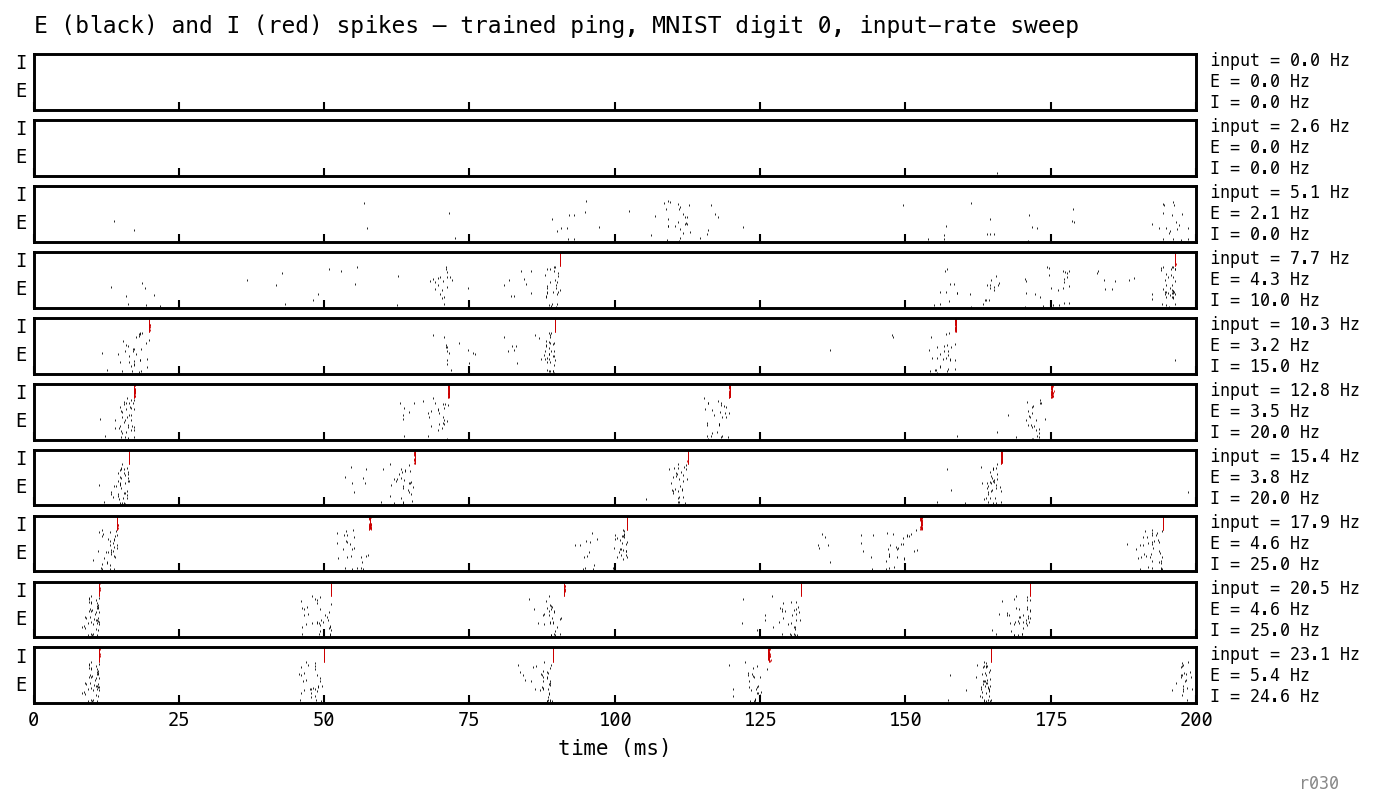
Trained PING (the nb025 baseline) replayed on MNIST digit 0 across ten input-rate values from 0 up to ≈ 23 Hz; each panel is one rate, E spikes (black) above I spikes (red), labelled with the input rate and the resulting per-cell E and I rates. Reading down the stack: at low drive the network is silent (E and I both 0); past a recruitment threshold (≈ 8 Hz input) the I-loop catches — the I rate jumps from 0 to ≈ 10 Hz and gamma bands appear in the E raster; above that the E rate stays pinned near 4–5 Hz while the I rate keeps climbing toward ≈ 25 Hz, and the cycle period barely moves with further drive. (The faint clustering in the just-engaged panels, where I is still 0, is feedforward input structure rather than PING — see the note below.)
At rate = 0 the network is silent. Above a recruitment threshold a few E units fire, the I loop catches, and the gamma cadence appears. Once the loop is locked the cycle period is invariant to further increases in input rate.
A subtlety in the just-engaged panels (e.g. input Hz, E = 1.7 Hz, I = 0): the E spikes look mildly clustered even though the I-loop hasn’t fired. This is feedforward — not PING. The Poisson input stream has natural temporal fluctuations, the digit-0 input is spatially correlated, and the trained has learned to be selective for digit-relevant pixels. So the cells that fire are the input-tuned subset, and they fire together during input-rich windows. The participation pattern is partially preserved from training; the gamma timing is not, because the loop is genuinely off.
To isolate the bare drive-response of each architecture, repeat the sweep with a spatially uniform input — every input channel firing at the same Poisson rate, with no digit structure — and compare PING against COBA on the same axes.
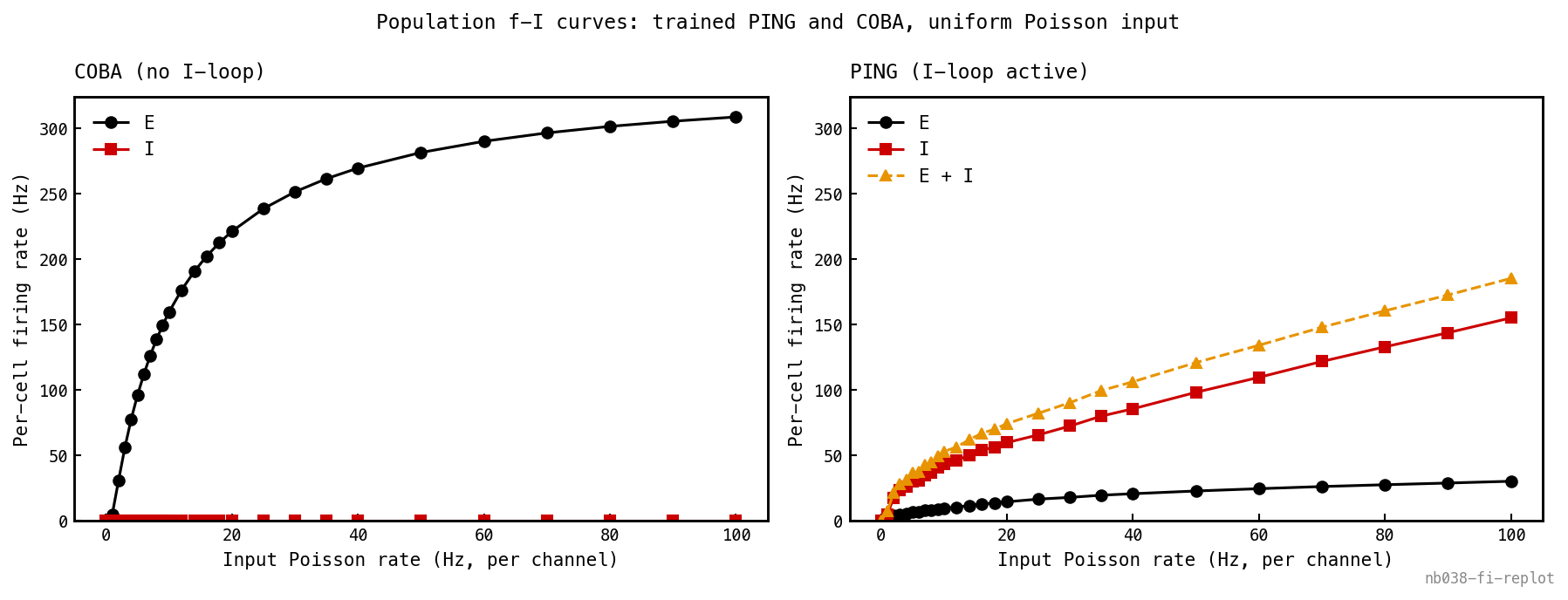
Population f-I curves — mean per-cell firing rate vs input Poisson rate — for trained COBA (left) and trained PING (right) under spatially uniform input (all 784 channels at the same rate, no digit structure), both panels on a shared y-axis so the compression is read on one scale. COBA (left): E (black) climbs steeply and saturates toward its refractory ceiling (≈ 310 Hz at 100 Hz input — every cell firing about as fast as it can), with I (red) flat at zero because the loop is disabled. PING (right): a recruitment cliff at low input where the I-loop engages, after which E is held ≈ 10× lower than COBA across the range — only ≈ 30 Hz at 100 Hz input where COBA runs to ≈ 310. The I curve (red) and the E+I total (amber) ride above the E curve: I fires harder than E to suppress it. This is the function of PING — dynamic-range compression, folding drive that would saturate E into a bounded gamma cadence.
Two qualitative responses to the same drive, on one shared y-axis. COBA (left) rises sharply across 1–10 Hz input then saturates toward its refractory ceiling ( Hz at 100 Hz input — every E cell firing essentially as fast as it can); the I rate stays at zero because the loop is off. PING (right) shows the recruitment cliff at low input and then holds E roughly an order of magnitude lower than COBA across the entire post-engagement range — Hz at 100 Hz input against COBA’s . The I curve sitting above the E curve at every input rate is the visual signature of the loop — I has to fire harder than E to suppress it. The I-loop is doing dynamic-range compression — pulling drive that would saturate the E population into a bounded gamma cadence.
Discussion
The signed-current picture (Figure 4c) is the load-bearing observation: stays non-negative throughout, but the current it drives is negative because — sign lives in the driving force, never in the conductance. The f-I curves make the function of the loop visible: PING is dynamic-range compression, not just rhythm. E rate stays an order of magnitude below COBA across two orders of magnitude of input, while COBA saturates near its refractory ceiling.
Next steps
- Sweep and independently and confirm the gamma peak tracks .
- Repeat the f-I curves with a sweep of refractory period and check whether COBA’s saturation point lines up with the predicted ceiling.